UNIDADE II - PATRIMÓNIO GENÉTICO
1.Património genético
1.1. Transmissão de características hereditárias
1.2. Organização e regulação do material genético
2.Alterações do material genético
2.2. Fundamentos da engenharia genética
1.Património genético
![]()
1.1. Transmissão de características hereditárias
Vídeo http://sci2.tv/#!/videos/1027 2’15’’
![]() Gregor Mendel
Gregor Mendel
Gregor Mendel nasce em 1822, na Áustria e ingressa num mosteiro em Brunn em 1943 na atual República Checa. Foi ordenado monge em 1847, estudou na universidade de Viena, matemática e ciências, o seu objetivo era ser professor mas foi mal sucedido nos exames. De volta ao mosteiro, onde passou o resto da sua vida, Mendel estudou abelhas e cultivou plantas, inclusive chegou a criar novos frutos (maças e peras). Entre 1856 e 65, Mendel realizou uma série de experiências que visavam as características hereditárias transmitidas de geração a geração. Em 1865 apresentou as suas leis de hereditariedade à Sociedade de História Natural de Brunn, deduzidas a partir das experiências que tinha feito em ervilhas. Pela falta de conhecimento da época poucos foram os que entenderam o trabalho de Mendel e só em 1900 foram redescobertas por 3 investigadores que trabalhavam independentemente. Mendel não viveu para ver o seu trabalho reconhecido e hoje assume uma grande importância na ciência, sendo considerado o “Pai da Genética”.
A escolha da planta
Mendel escolheu a ervilheira porque esta tem um ciclo reprodutivo curto, produz muitas sementes, os componentes envolvidos na reprodução sexuada ficam encerrados no interior da flor, protegidas pelas pétalas, o que favorece a autopolinização e consequentemente a autofecundação, formando descendentes com as mesmas caraterísticas da progenitora, e ainda pela existência de uma grande variedade de caraterísticas de fácil comparação. Por exemplo: a cor das ervilheiras, umas púrpuras e outras brancas, sementes lisas e rugosas, etc.
Os experimentos de Mendel

http://www.sobiologia.com.br/conteudos/Genetica/leismendel.php
No início da experiência Mendel produziu linhagens puras de ervilhas, através da autopolinização, para determinada característica que pretendia estudar. Por exemplo: a autopolinização das ervilheiras de cor púrpura, davam sempre descendência de cor púrpura, e para as brancas a mesma coisa. A autopolinização assegura a pureza das linhagens.
Mendel estudou sete características: cor da flor, posição da flor no caule, cor da semente, aspeto externo da semente, forma da vagem, cor da vagem e altura da planta.
Após a obtenção de linhagens puras, Mendel utilizou uma diferente reprodução, polinização cruzada. Cruzou linhagens puras entre si, e estas são a geração parental (P), ou seja os progenitores. Por exemplo, Mendel, para a característica da cor das sementes, cortou os estames de uma planta de vagem amarela e depositou no estigma de uma outra flor, com cor diferente de semente, cor verde.
Depois de repetir este procedimento várias vezes, Mendel verificou que os descendentes híbridos (cruzamento de linhagens diferentes), a que Mendel designou por primeira geração filial (F1), eram todas de cor amarela, o que Mendel conclui que esta característica, cor da semente amarela, era dominante, e a cor verde era recessiva, não apareceu uma única vez.
O procedimento seguinte foi cruzar os descendentes híbridos da primeira geração (F1) até surgirem as plantas e as flores. A reprodução foi natural, auto fertilizaram-se e já obteve sementes verdes em menor quantidade que as amarelas, na proporção 1:3, respetivamente, na geração F2.

http://www.qieducacao.com/2011/05/genetica-1-lei-de-mendel.html
Com estes resultados Mendel pode concluir Concluiu que a cor verde das sementes não se tinha manifestado na primeira geração mas que não tinha “desaparecido” nas sementes da geração F1. Concluiu que a cor verde era um caráter recessivo e a cor amarela era a dominante. Mendel deduziu que a cor das sementes era determinada por dois fatores que fazia surgir uma cor, amarela ou verde.
Na representação destes fatores, Mendel escolheu a letra maiúscula para o caracter dominante e a minúscula para o recessivo.
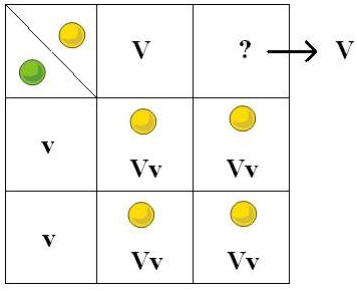
Assim, para este experimento teríamos o v (para o verde recessivo) e o V (para o amarelo dominante).
|
Semente amarela pura |
Semente amarela híbrida |
Semente verde pura |
|
VV |
Vv |
vv |
Como explicar o desaparecimento da cor verde na geração F1 e o seu reaparecimento na geração F2.
A resposta surgiu a partir do conhecimento de que cada um dos fatores se separava durante a formação dos gametas. O material hereditário passa de uma geração para a outra.

http://www.sobiologia.com.br/conteudos/Genetica/leismendel.php
Na segunda geração (F2) para cada 3 sementes amarelas, Mendel obteve apenas uma semente verde, o mesmo se passou para todas as outras 6 características estudadas por Mendel. A proporção foi sempre de 3 dominantes para uma recessiva. Esta comprovação da existência de dominância e recessividade, levou a Mendel a formular a sua primeira lei.
![]() 1ª
lei de Mendel da segregação dos fatores
que diz:
“ Cada característica é determinada por dois
fatores que se separam na formação dos gametas, onde ocorrem em dose simples”,
isto é, para cada gameta masculino ou feminino encaminha-se apenas um fator.
1ª
lei de Mendel da segregação dos fatores
que diz:
“ Cada característica é determinada por dois
fatores que se separam na formação dos gametas, onde ocorrem em dose simples”,
isto é, para cada gameta masculino ou feminino encaminha-se apenas um fator.
Embora Mendel tivesse sido muito inovador para a época, Mendel não sabia onde estavam localizados esses fatores hereditários e como é que eles se segregavam. Só em 1900, com a redescoberta de Mendel, é que se levantou esta questão. Walter Sutton, em 1902, ao estudar a formação dos gametas dos gafanhotos, lançou a hipótese que os pares de fatores hereditários estavam localizados nos cromossomas homólogos, e com a meiose, a separação dos homólogos levava à segregação dos fatores.
Hoje sabemos que os fatores a que Mendel se referiu são os genes (do grego genos, originar, provir), e que realmente estão localizados nos cromossomos, como Sutton propôs. Gene são fragmentos funcionais de DNA cuja atividade pode originar o aparecimento de um fenótipo* observável. As diferentes formas sob as quais um gene pode se apresentar, os tais fatores a que Mendel se referia, são denominadas de genes alelos, ou simplesmente de alelos sendo cada caracter determinado por duas formas alternativas para o mesmo gene e estes encontram-se no mesmo local em cromossomas homólogos, Locus.

Por exemplo: a cor amarela e a cor verde da semente de ervilha, por exemplo, são determinadas por dois alelos, isto é, duas diferentes formas do gene para cor da semente.
![]() Cruzamentos teste
Cruzamentos teste
Hoje em dia para o estudo do genótipo dos indivíduos pode ser feito de uma forma rápida mas muito dispendiosa, que é analisar o DNA. Por este motivo ainda se recorre ao método tradicional em que se promove o cruzamento de homozigóticos recessivos, para uma determinada característica, e se analisa a respetiva descendência.
Quando não se sabe se o indivíduo é homozigótico dominante ou heterozigótico, realizam-se cruzamentos entre este indivíduo com um que saibamos que é recessivo e a resposta estará nos descendentes. Se a descendência for recessiva, significa que o progenitor é também recessivo. Se todos os descendentes tiverem um fenótipo dominante, quer dizer que o progenitor desconhecido é um homozigótico.

http://www.infoescola.com/biologia/termos-usados-em-genetica/
Ao cruzar um progenitor homozigótico recessivo (verde) com o que pretendemos determinar amarelo, obteve-se na geração F1 descendentes verdes, isso indica que o amarelo testado era heterozigoto Vv, pois quando cruzamos um heterozigoto com um recessivo, 50% da prole é dominante (heterozigótica) e 50% é recessiva.
http://www.infoescola.com/biologia/termos-usados-em-genetica/
Aqui, como toda a geração F1 é amarela, quer dizer que o progenitor amarelo é homozigótico dominante. Quando cruzamos dois indivíduos homozigotos (puros), sendo um dominante e um recessivo, 100% da prole é heterozigótica.
Retrocruzamento: é um processo muito parecido com o cruzamento teste, pois utiliza a mesma técnica, porém o indivíduo recessivo utilizado no teste é de algum ancestral do indivíduo com genótipo dominante desconhecido.
![]() Rescrevendo as leis de Mendel
com mais conhecimentos de genética.
Rescrevendo as leis de Mendel
com mais conhecimentos de genética.
1ª lei de Mendel – Os genes alelos separam-se na formação de gametas e a probabilidade de um gameta transportar um dos alelos é igual a do outro gameta transportar o outro alelo.
2ª lei de Mendel – Na gametogénese a segregação de um determinado alelo de um gene é independente da segregação de um outro alelo de um outro gene.
![]() Os
conceitos de fenótipo e genótipo
Os
conceitos de fenótipo e genótipo
Estes conceitos forma criados pelo dinamarquês Wilhelm L. Johannsen, no principio do século XX.
Fenótipo
O termo “fenótipo” (do grego pheno, evidente, brilhante, e typos, característico) às características apresentadas por um indivíduo, sejam elas morfológicas, fisiológicas e comportamentais. Também fazem parte do fenótipo características microscópicas e de natureza bioquímica, que necessitam de testes especiais para a sua identificação.
A cor de olhos, cor de pele, textura de cabelo, etc, são características visíveis, no entanto, o tipo de sangue, a sequência de aminoácidos de uma proteína são características fenotípicas apenas identificadas por testes.
O fenótipo de um indivíduo sofre alterações ao longo da vida e o próprio ambiente pode contribuir para esta modificação. O nosso corpo modifica-se, o nosso cabelo fica branco, a nossa pele exposta à luz escurece, etc..
Genótipo
O termo “genótipo” (do grego genos, originar, provir, e typos, característica), diz respeito à constituição genética do indivíduo, ou seja, está escrito nos genes que possui.
Por exemplo se uma pessoa tem olhos azuis, dizemos que ela é homozigótica em relação à cor de olhos (aa), ou heterozigótica (aC) se tiver olhos castanhos, em que a cor castanha é dominante à cor azul.
O genótipo é inferido a partir do estudo do fenótipo, através do sequenciamento do genoma (alto custo) ou através da análise da família, através da observação.
![]() Mecanismos de transmissão hereditária de dois pares
de genes
Mecanismos de transmissão hereditária de dois pares
de genes
Mendel analisou a transmissão hereditária de dois pares de genes, o bi-hibridismo, selecionando linhas puras de ervilheiras que diferenciavam-se entre si por duas características: a cor e forma da semente.
O cruzamento de linhas puras origina híbridos, se são de duas características os descendentes chama-se bi-híbridos.
Mendel cruzou ervilheiras produtoras de sementes amarelas e lisas (ambas características dominantes - AALL) com ervilheiras produtoras de sementes verdes e rugosas (ambas recessivas – aall). Os híbridos resultantes (F1) são, obrigatoriamente, heterozigóticos, AaLl, e fenotipicamente apresentam-se amarelas e lisas, pois contém os dois alelos dominantes.
No cruzamento seguinte, F2, através da autopolinização a geração que surge, 16 indivíduos, é constituída por 4 fenótipos na seguinte proporção:
-9 Indivíduos com sementes amarelas e lisas em 16;
-3 Indivíduos com sementes amarelas e rugosas em 16;
-3 Indivíduos com sementes verdes e lisas em 16;
-1 Indivíduo com sementes verdes e rugosas em 16.
Posto isto, verifica-se que cada par de alelos funciona da mesma forma do moniidibrismo.
Fazendo as contas:
Por exemplo para a cor das sementes, existem 12 sementes em 16 de cor amarela para 3 verdes, a proporção continua a ser 3:1. Este facto está de acordo com a Primeira lei de Mendel.
Se utilizarmos as leis das probabilidades o resultado é o mesmo.
Vejamos: a probabilidade de um gameta receber uma das cores é ½, ou seja, 50% probabilidade para receber a cor amarela e 50% de probabilidade de receber a cor verde. O mesmo acontece para a característica da textura da semente, 50% de probabilidade de ser lisa e 50% de probabilidade de ser rugosa.
Aplicando os princípios das probabilidades temos:
(1/2 A +1/2 a) x (1/2 L + 1/2 l) =
1/4 AL + 1/4 aL + 1/4 Al + 1/4 al
Atendendo à dominância e recessividade teremos os mesmo resultados de Mendel:
-9 Indivíduos com sementes amarelas e lisas em 16;
-3 Indivíduos com sementes amarelas e rugosas em 16;
-3 Indivíduos com sementes verdes e lisas em 16;
-1Indivíduo com sementes verdes e rugosas em 16.
Conclui-se que os resultados experimentais obtidos por Mendel não se afastam do modelo teórico das leis de probabilidades.
![]() Cruzamentos-teste em di-hibrismo
Cruzamentos-teste em di-hibrismo
Existe um paralelismo entre este tipo de testes e o que faz para o monibrismo, ou seja, procede-se da mesma forma, no entanto
neste caso tem de se utilizar um ser duplamente recessivo para os alelos dominantes expressos no fenótipo do individuo que se pretende testar.
Exemplo:
Pretende-se conhecer o genótipo das ervilheiras com fenótipo que expressou a semente amarela e lisas (características dominantes A?,L?), e ir-se-á cruzar estas com um fenótipo recessivo, ou seja, sementes verdes e rugosas (aall).
Duas hipóteses:

Da Silva, A. D., Santos, M. E., Mesquita, A. F., Baldaia, L., Félix, J. M. Terra, Universo da Vida - Biologia. 12º ano, Porto Editora, Porto, Portugal, 2012 - páginas72 a 76
![]() Exceções às leis de Mendel
Exceções às leis de Mendel
Dominância Incompleta – situações nas quais o fenótipo dos heterozigóticos é intermédio entre os dois homozigóticos, sem ocorrer mistura dos alelos, pois aparecem homozigóticos em F2.
Alelos Múltiplos (Polialelismo) – existência de mais de dois alelos num dado gene.
Podem surgir alterações num alelo de um indivíduo, que, posteriormente, o pode transmitir à sua descendência, provocando o aparecimento de mais de dois alelos para um dado gene numa população. Todavia, cada indivíduo apenas possui dois alelos, herdados dos seus progenitores.
Codominância – nenhum alelo exerce dominância sobre o outro e ambos se expressam fenotipicamente.
Alelos letais – causam a morte pré ou pós-natal, ou então produzem uma deformidade significante. A combinação letal (em homozigotia recessiva) modifica a proporção dos fenótipos dos sobreviventes (2:1).
Codominância
Nem todas as características são herdadas como a cor da semente da ervilha, em que o gene para a cor amarela domina sobre o gene para cor verde. Muito frequentemente a combinação dos genes alelos diferentes produz um fenótipo intermediário, é chamada de dominância incompleta ou parcial.
A codominância é quando os dois alelos do genótipo se expressam no fenótipo, simultaneamente.
Um exemplo de Codominância são a raça bovina Shorthorn.

O cruzamento entre indivíduos linhas puras de cor vermelha com indivíduos linhas puras de cor branca, origina descendentes que possuem uma mistura de pelos vermelhos e pelos brancos, cujo efeito é uma coloração cinzento-avermelhada. Cada um dos alelos expressa-se de forma independente, pelo a pelo. Diz-se que existe um situação de codominância.
Dominância Incompleta
O terceiro fénotipo é intermédio dos fenótipos dos dois progenitores.


A inexistência de uma verdadeira relação dominância/recessividade entre os alelos responsáveis pela cor das flores, ou seja, não há dominância total do alelo vermelho sobre o alelo branco. Diz-se, por isso, que existe uma dominância incompleta.
Gene letal
Gene letal é um gene que, quando presente, provoca a morte pré ou pós nascimento, ou produz uma deformidade tão grave que levará o indivíduo à morte.
Os alelos letais podem ser tanto dominantes ou recessivo, se forem dominantes levam à morte quando aparecerem em homozigóticos ou heterozigóticos dominantes, se forem recessivos, levando à morte quando em homozigóticos recessivos.
Quando a proporção dos descendentes não obedecer a proporção esperada segundos as Leis de Mendel é de suspeitar que existam genes letais no DNA.
No principio do século, Lucien Cuenot, geneticista francês, estudou os camundongos e verificou que os resultados se afastavam dos que tinha previsto Mendel. Cuenot tinha por objetivos obter um de cor amarela que fosse de linha pura (homozigóticos) mas sem sucesso, pois todos os ratos que obteve de cor amarela eram heterozigóticos, os homozigóticos não chegavam a nascer.
Exemplo: retirado de http://www.uel.br/pessoal/rogerio/genetica/respostas/pratica_08.html
Gene letal em camundongos

- Nestes animais, o gene K/k está envolvido em duas vias metabólicas diferentes, uma delas relacionada com a coloração da pelagem e uma outra com o desenvolvimento embrionário:
Em relação à coloração, o alelo que determina a cor amarela é dominante sobre a aguti (castanho avermelhado).
Quanto a sobrevivência ou morte, esses mesmos alelos possuem um efeito contrário: dominante para a sobrevivência e recessivo para a letalidade.
Logo, e observando a figura, não é possível obter uma linhagem pura de camundongos de coloração amarela (KK), tendo em vista que eles morrem durante o desenvolvimento embrionário.
Polialelia ou alelos múltiplos
Significa que existem três ou mais tipos de alelos distintos para os mesmos locus cromossómicos. Cada indivíduo tem apenas um par desses genes, mas as combinações possíveis entre elas são várias.
Resulta de mutações sucessivas ocorridas nos genes de um determinado locus. Cada gene mutante que surge estabelece condições para o aparecimento de mais uma nova expressão fenotípica daquele caráter, naquela espécie.
Exemplo: Sistema sanguíneo ABO
Sistema proposto, em 1900, pelo austríaco Landsteiner, que estabelecia para o homem 4 quatro grupos sanguíneos, A, B AB e O, considerando a relação entre os pares dos alelos: IA, IB e i, em quatro grupos: grupo A, grupo B, grupo AB e grupo O.
As nossas hemácias podem apresentar na membrana
substâncias designadas por aglutinogénios, sintetizadas pelos alelos IA ou
IB sendo: aglutinogénio A ou aglutinogénio B, aglutinogénio A e
aglutinogénio B e também a substância química aglutinina contida no plasma das
hemácias: Anti-A, Anti-B ou a ausência dessas.
Na relação alélica existente, o alelo i é recessivo aos seus alelos IA e
IB. Se o indivíduo é homozigótico recessivo (genótipo ii) este grupo
sanguíneo pertence ao grupo O.
Se forem heterozigóticos, os alelos IA e IB, ambos
manifestam seu caráter dominante, e o indivíduo será do grupo sanguíneo AB
(genótipo IA IB).
Um indivíduo pertencerá ao grupo sanguíneo A, se enquadrado em duas situações:
quando é homozigótico dominante IA IA, ou em
heterozigótico do alelo dominante IA com o recessivo i, apresentando genótipo IA i.
Da mesma forma para o grupo sanguíneo B: quando é homozigótico dominante IB IB,
ou em heterozigótico do alelo dominante IB com o recessivo i,
apresentando genótipo IB i.
Possibilidades entre os alelos para
determinação do sistema ABO.
|
Grupo sanguíneo |
Genótipo |
Aglutinogénio |
Aglutinina |
|
A |
IA IA ou IA i |
A |
Anti-B |
|
B |
IB IB ou IB i |
B |
Anti-A |
|
AB |
IA IB |
AB |
Ausência |
|
O |
ii |
Ausência |
Anti-A e Anti-B |

adaptado de http://grado9.wordpress.com/2011/02/07/genetica/
![]() Interações génicas
Interações génicas
Ocorrem quando as características herdadas são controladas por mais que um gene que se pode localizar num diferente locus dentro do mesmo cromossoma ou mesmo em cromossomas diferentes.
Um exemplo disto é a cor dos olhos do humano.
A cor da íris é determinada por dois tipos de melanina produzidos pelos melanócitos, ou células produtoras de pigmentos, na íris. Sabe-se hoje que a cor dos olhos, a grande quantidade de variações existentes na cor, não são resultado de apenas um tipo de gene que determina a cor, mas trata-se de uma herança poligénica (mais que um gene), um tipo de variação continua em que os alelos de vários genes dão a cloração aos olhos. Existem proteínas que comandam a proporção de melanina depositada na íris, outros genes produzem manchas, raios e padrões de difusão na cor de olhos.
Os olhos castanhos são os que têm mais melanina, os azuis os que têm menos e os verdes apenas (2% da população tem olhos verdes), têm mais que as azuis e menos que os castanhos.
A interação entre genes (interações génicas) são muito frequentes na natureza: cor de pele, estatura, cor de olhos, são algumas destas interações.
Ver página 84 – cor de pelo nos cães labradores.
![]() Teoria Cromossómica da Hereditariedade – Em cada
cromossoma existe uma sequência de genes.
Teoria Cromossómica da Hereditariedade – Em cada
cromossoma existe uma sequência de genes.
Esta teoria é defendida por Walter Sutton e Theodor Boveri (1902) e permite explicar, através de processos celulares, as conclusões obtidas por Mendel.
Pressupostos:
-Os genes estão localizados em cromossomas;
-Os cromossomas formam pares de homólogos que possuem no mesmo locus alelos para o mesmo caracter;
-Em cada par de cromossomas, um tem origem materna e o outro de origem paterna;
-Durante a meiose ocorre disjunção dos homólogos que são transmitidos aos gâmetas, promovendo a segregação dos alelos;
-A segregação dos alelos localizados em cromossomas diferentes é independente;
-Através da fecundação, há formação do ovo, em que cada gene está representado por dois alelos, localizados em loci (plural de locus) correspondentes de cromossomas homólogos.
![]() Hereditariedade
ligada aos cromossomas sexuais
Hereditariedade
ligada aos cromossomas sexuais
No cariótipo de um individuo (conjunto de cromossomas existente no núcleo de cada uma das células somáticas) distinguem-se os cromossomas que determinam o sexo, são os cromossomas sexuais ou HETEROSSOMAS, dos restantes cromossomas, os AUTOSSOMAS. Na reprodução sexuada cada individuo é portador de DNA transmitido pelos seus progenitores, parte é de origem materna e outra de origem paterna expressos nos cromossomas homólogos e em que o gene de uma determinada característica se encontra no mesmo locus no cromossoma homólogo.
Normalmente nos seres diplontes os cromossomas que determinam o sexo são diferentes entre si, por exemplo o sexo feminino na mulher é determinado pelo cromossoma 23 e tem dois cromossomas iguais (XX) e o homem diferentes (XY), ou seja no homem este par de cromossomas não é totalmente homólogo.
Trabalhos de Morgan – Hereditariedade ligada ao sexo e Ligação Fatorial
Thomas H. Morgan, foi um embriologista da Universidade de Colúmbia, desenvolveu em 1910 estudos genéticos com a mosca da fruta Drosophila melagaster. Foi prémio Nobel em 1933, pelo seu contributo para o avanço da genética, provando que os cromossomas são portadores de genes.

http://pt.wikipedia.org/wiki/Drosophila
Morgan iniciou os seus estudos estudando a hereditariedade da cor dos olhos da Drosophila. A cor de olhos da mosca da fruta selvagem mais frequente é cor vermelha, no entanto, outras moscas tinham olhos de cor branca (mutantes). Começou então a cruzar fêmeas de olhos vermelhos com machos de olhos brancos. A escolha da Drosophila como excelente “material” biológico a estudar está relacionado por: ter dimensões reduzidas; ter um ciclo de vida muito curto e que produz um número elevado de descendentes; distinguir-se bem a fémea do macho (dimorfismo sexual), ter um número reduzido de cromossomas; possuir uma diversidade de características controláveis e a cultura e manipulação em laboratório serem relativamente fáceis.
A Drosophila tem um cariótipo constituído por apenas 8 cromossomas (4 pares), logo tem necessariamente mais que um gene em cada cromossoma.

Morgan, cruzou então as fêmeas de olhos vermelhos com os machos de olhos brancos e obteve na primeira geração (F1), descendentes apenas com cor vermelha, o que demonstra que o alelo da cor branca é recessiva. Até aqui, tudo de acordo com os estudos de Mendel, no entanto verificou-se que os olhos brancos eram apenas se manifestavam nos machos e embora a cor vermelha se manifestasse também nos machos, a proporção relativa às fêmeas era muito menor (duas fêmeas para dois machos).
Morgan sabia que dos 4 pares de cromossomas da mosca, um determinaria o sexo. Na fêmea estes cromossomas eram de morfologia igual XX e no macho diferente XY, em que Y é mais pequeno e com menos genes que X.
Na formação dos gametas, na meiose, nas fêmeas, os gametas serão sempre iguais, dado que provém de dois cromossomas XX, mas nos machos um gameta é X e outro é Y. Quando ocorre a fecundação, o zigoto trás consigo informação específica relativamente ao sexo: se os dois gametas forem X, então forma-se uma fêmea se um for X e outro Y, forma-se um macho. Posto isto, Morgan deduziu: Se a cor dos olhos da mosca-da-fruta se encontra no cromossoma X, este manifestar-se-á sempre no fenótipo dos machos, pois não existe outro alelo no cromossoma Y.

Morgan coloca, então, a hipótese que o alelo que determina a cor dos olhos da mosca-da-fruta se encontra no cromossoma X (hereditariedade ligada ao sexo - características cujos genes responsáveis pela sua manifestação se localizam num cromossoma sexual dizem-se características ligadas ao sexo).
O seguinte procedimento foi fazer o cruzamento reciproco, ou seja cruzar um macho de olhos vermelhos e uma fêmea de olhos brancos, e o resultado foi: 25% de fêmeas com olhos vermelhos; 25% de fêmeas com olhos brancos; 25 % de machos com olhos vermelhos e 25% de machos de olhos brancos.
Ao interpretar os dados dos dois cruzamentos segundo a hipótese de Morgan ( alelo que determina o cor está no cromossoma X) permitiu explicar os resultados obtidos. E Morgan concluiu que os genes se encontravam nos cromossomas e alguns deles nos cromossomas sexuais.
Ligação fatorial
Cada cromossoma tem uma série de genes dispostos em linha – genes ligados fatorialmente ou em Linkage, mas os genes presentes no mesmo cromossoma não se comportam como uma unidade indissociável, pois durante a meiose (prófase I), na formação de gâmetas, podem ocorrer fenómenos de crossing-over.

Morgan cruzou moscas selvagens de corpo cinza e asas longas com mutantes de corpo preto e asas curtas (chamadas de asas vestigiais). Todos os descendentes de F1 apresentavam corpo cinza e asas longas, em que o gene que condiciona corpo cinza (P) domina o que condiciona o corpo preto (p), assim como o gene para asas longas (V) é dominante sobre o (v) que condiciona surgimento de asas vestigiais.
A seguir Morgan cruzou descendentes de F1 com duplo-recessivos. O objetivo era que os resultados dos cruzamentos-teste revelassem se os genes estavam localizados em cromossomas diferentes (segregação-independente) ou no mesmo cromossoma (linkage). Porém, nenhum dos resultados esperados foi obtido. A separação e a contagem dos descendentes de F2 revelou o seguinte resultado:
41,5% de moscas com o corpo cinza e asas longas;
41,5% de moscas com o corpo preto e asas vestigiais;
8,5% de moscas com o corpo preto e asas longas;
8,5% de moscas com o corpo cinza e asas vestigiais.
Ao analisar esse resultado, Morgan convenceu-se de que os genes P e V localizavam-se no mesmo cromossomo. Se estivessem localizados em cromossomos diferentes, a proporção esperada seria outra (1: 1: 1: 1). No entanto, restava a dúvida: como explicar a ocorrência dos fenótipos corpo cinza/asas vestigiais e corpo preto/asas longas?
A resposta não foi difícil de ser obtida, estava relacionada com os fenótipos corpo cinza / asas vestigiais e corpo negro /asas longas, eram recombinantes e pela ocorrência de crossing-over na meiose.

Na Prófase I da meiose pode ocorrer crossing-over entre os dois cromossomas homólogos. Estas regiões do cromossoma podem ser trocadas reciprocamente, tornando-se recombinantes. Assim, nem todos os descendentes possuirão os fenótipos parentais, surgindo em seu lugar fenótipos recombinantes.
As frequências de recombinação são tanto maiores quanto mais afastados estiverem os dois loci, uma vez que tal facilita a separação e recombinação durante a meiose.
Quando comparamos o comportamento de pares de genes para duas características para a segunda lei de Mendel com a ocorrência de linkage e crossing-over em um cruzamento genérico do tipo AaBb X aabb, verificamos que em todos os casos resultam quatro fenótipos diferentes:
Dominante/dominante
Dominante/recessivo
Recessivo/dominante
Recessivo/recessivo.
A diferença em cada caso está nas proporções obtidas. No caso da 2ª lei de Mendel, haverá 25% de cada fenótipo. No linkage com crossing, todavia, os dois fenótipos parentais surgirão com frequência maior do que as frequências dos recombinantes.
A explicação para isso reside no fato de, durante a meiose a permuta não ocorrer em todas as células, sendo, na verdade, um evento relativamente raro.
Frequentemente, nos vários cruzamentos realizados do tipo AaBb X aabb, Morgan obteve os dois fenótipos parentais (AaBb e aabb), na proporção de 50% cada. Para explicar esse resultado, ele sugeriu a hipótese que os genes ligados ficam tão próximos um do outro que dificultam a ocorrência de crossing over entre eles. Assim, por exemplo, o gene que determina a cor preta do corpo da drosófila e o gene que condiciona a cor vermelha dos olhos ficam tão próximos que entre eles não ocorre permuta. Nesse caso se fizermos um cruzamento teste entre o duplo-heterozidoto e o duplo-recessivo, teremos nos descendentes apenas dois tipos de fenótipos, que serão correspondentes aos tipos parentais.
Na Hereditariedade ligada ao sexo as fêmeas podem ser heterozigóticas para genes localizados no cromossoma X, contudo, os machos serão sempre hemizigóticos* no que respeita aos genes localizados no cromossoma X, uma vez que só possuem um alelo para estes genes que é sempre expresso.
Hemizigótico - único alelo que está localizado no cromossoma X, dado que o Y não contém esse alelo.
![]() Se
a característica estiver localizada em cromossomas sexuais, em cruzamentos
recíprocos, pode originar resultados diferentes entre machos e fêmeas.
Se
a característica estiver localizada em cromossomas sexuais, em cruzamentos
recíprocos, pode originar resultados diferentes entre machos e fêmeas.
![]() Da
análise de fenótipos
recessivos ligados
ao cromossoma
X podemos concluir
que:
Da
análise de fenótipos
recessivos ligados
ao cromossoma
X podemos concluir
que:
-O fenótipo surge com muito maior frequência em machos do que em fêmeas, pois estas necessitariam dos dois alelos recessivos;
-Um macho com a doença apenas as pode transmitir às suas filhas, pois os filhos recebem o cromossoma Y do pai;
-As filhas que recebem um cromossoma X com o alelo para a doença são heterozigóticas portadoras, apresentando-se fenotipicamente normais. Neste caso, podem transmitir a doença à sua descendência.
- pode haver gerações que não manifestam o fenótipo.
Quando o alelo responsável pela doença se localiza no cromossoma Y todos os filhos de pais com a anomalia apresentam a doença.
![]() Hereditariedade humana
Hereditariedade humana
A investigação nos humanos é difícil, ao contrário do que já referimos nas moscas-da-fruta, nós temos um grande número de cromossomas, o nosso ciclo de vida é longo, temos poucos descendentes e obviamente que não podemos fazer cruzamentos experimentais.
Esta investigação pode ser feita de uma forma tradicional e menos onerosa através de árvores genealógicas ou heredogramas de famílias que nos poderão permitir perceber a hereditariedade ao longo de várias gerações na transmissão de determinados caracteres.
Através da análise de uma árvore genealógica é possível determinar se um alelo é recessivo ou dominante e ainda a sua localização em autossomas (hereditariedade autossómica) ou em heterossomas (hereditariedade ligada ao sexo).
Para que todos possam entender as árvores, em todas as situações e em qualquer parte do mundo, foi necessário criar uma simbologia.

Interpretação dos Heredogramas
“A análise pode permitir se determinar o padrão de herança de uma certa característica (se é autossómica, se é dominante ou recessiva, etc.). Permite, ainda, descobrir o genótipo das pessoas envolvidas, se não de todas, pelo menos de parte delas. Quando um dos membros de uma genealogia manifesta um fenótipo dominante, e não conseguimos determinar se ele é homozigoto dominante ou heterozigoto, habitualmente o seu genótipo é indicado como A_, B_ou C_, por exemplo.
A primeira informação que se procura obter, na análise de um heredograma, é se o caráter em questão é condicionado por um gene dominante ou recessivo. Para isso, devemos procurar, no heredograma, casais que são fenotipicamente iguais e tiveram um ou mais filhos diferentes deles. Se a característica permaneceu oculta no casal, e se manifestou no filho, só pode ser determinada por um gene recessivo. Pais fenotipicamente iguais, com um filho diferente deles, indicam que o caráter presente no filho é recessivo!
Uma vez que se descobriu qual é o gene dominante e qual é o recessivo, vamos agora localizar os homozigotos recessivos, porque todos eles manifestam o caráter recessivo. Depois disso, podemos começar a descobrir os genótipos das outras pessoas. Devemos nos lembrar de duas coisas:
1ª) Em um par de genes alelos, um veio do pai e o outro veio da mãe. Se um indivíduo é homozigoto recessivo, ele deve ter recebido um gene recessivo de cada ancestral.
2ª) Se um indivíduo é homozigoto recessivo, ele envia o gene recessivo para todos os seus filhos. Dessa forma, como em um “quebra-cabeças”, os outros genótipos vão sendo descobertos. Todos os genótipos devem ser indicados, mesmo que na sua forma parcial (A_, por exemplo).”

Exemplo de uma árvore genealógica
Retirado de: http://www.sobiologia.com.br/conteudos/Genetica/leismendel5.php
Com o avanço da genética desenvolveram – se técnicas de genética molecular que permitem análise direta de genes e cujo objetivo servem para diagnóstico e despiste de doenças genéticas. Devido ao custo elevado, ainda são utilizadas as árvores genealógicas que depois de analisadas nos podem fornecer dados da transmissão de um determinado gene e posteriormente, caso seja considerado algo nocivo e que se queira avaliar o risco, poder-se-ão fazer a análise da genética molecular.
As genealogias humanas nem sempre mostraram as proporções de Mendel, isto deve-se ao facto de o homem ter um número reduzido de descendentes.
As genealogias podem representar características que não apresentam dominância, como é o caso da textura dos cabelos, que é um caso de herança sem dominância.

Nesta árvore genealógica o indivíduo 8 (F2) é portador de uma característica, e os seus pais não (F1). Em genética, indivíduos com características genéticas diferentes dos pais são recessivos, e os pais automaticamente são heterozigotos. Se os pais fossem recessivos, também estariam destacados com cor diferente. Pais recessivos só podem ter filhos recessivos, pois não têm genes dominantes para passar à descendência. Os filhos são recessivos porque os pais heterozigotos são portadores do gene recessivo.

Fonte : http://www.infoescola.com/genetica/genealogia/
HEREDITARIEDADE AUTOSSÓMICA
Doenças genéticas por genes autossómicos recessivas
A Fenilcetorúnia (PKU) é uma doença genética rara que pode ser diagnosticada e prevenida logo à nascença, através do teste do pezinho. É uma doença é autossómica recessiva e afeta aproximadamente um em cada dez mil indivíduos da população caucasiana. Esta doença deve-se a uma mutação no gene fenilalanina hidroxilase que é uma enzima que catalisa a conversão da fenilalanina (aminoácido), presente nos alimentos, em tirosina (importante para a síntese da melanina), ao nível do fígado. Esta enzima, nas pessoas normais, vem de um gene presente no cromossoma 12, a pessoa com a mutação, possui em vez desta enzima, uma não funcional. Os homozigóticos recessivos, em relação a esta mutação, não catalizam o aminoácido em tirosina mas sim num ácido, o ácido fenilpirúvico. Este em conjunto com a fenilalina no sangue faz com que o desenvolvimento do cérebro da criança portadora seja afetado originando perturbações motoras e convulsões
Pelo facto dos progenitores poderem não apresentar este fenótipo, ou seja progenitores normais podem ter filhos e filhas com esta doença, leva a concluir que a doença é determinada por um alelo autossomático recessivo porque: os homens e as mulheres são igualmente afetadas, por isso não vem do sexo; a maioria dos afetados provém de progenitores normais; os heterozigóticos apresentam um fenótipo normal e dois progenitores afetados terão toda a sua descendência afetada.
Com os benefícios da genética, hoje, uma criança com esta doença, precocemente diagnosticada, poderá ter uma vida normal.

http://illustratedmedicine.blogspot.pt/2010_09_01_archive.html
Doenças genéticas por genes autossómicos dominantes
A doença de Huntington
Trata-se de doença hereditária rara, cerca de 3 a 7 casos em 100 mil habitantes, causada por uma mutação genética no cromossoma 4. É um distúrbio neurológico hereditário que causa movimentos do corpo anormais, com falta de coordenação, afetando o cérebro e aspetos da personalidade que se manifesta entre os 35 e os 45 anos.
Trata-se de doença autossómica dominante, então se um dos progenitores tem Huntington, a descendência tem 50% de chances de também desenvolver a doença. Se o descendente não herdar o gene da doença, não a desenvolverá nem a transmitirá à geração seguinte.
Por ser uma doença genética, atualmente não tem cura. No entanto, os sintomas podem ser minimizados com a administração de medicação.
![]() Hereditariedade ligada ao sexo
Hereditariedade ligada ao sexo
Daltonismo
O daltonismo é provocado por genes recessivos localizados no cromossomo X (sem alelos no Y), o problema ocorre muito mais frequentemente nos homens que nas mulheres. Estima-se que 10% da população masculina seja portadora do distúrbio, embora apenas 0,3 % das mulheres sejam atingidas.
DD -Mulher com visão normal-Homozigótica não portadora do gene anômalo (DD, normal)
Dd- Mulher com visão normal-Heterozigótica portadora do gene anômalo (Dd, normal)
dd- Mulher daltónica-Homozigótica recessiva (dd, daltónica)
D- Homem com visão normal-Homozigótico dominante (D, normal)
d- Homem daltónico-Homozigótico recessivo (d, daltónico)
É menos provável que a mulher seja daltónica do que o homem uma vez que para que a mulher seja daltónica necessita que os seus dois alelos sejam dd e no homem, como só possui um cromossoma X, basta aparecer neste o gene recessivo (d).
Se a mãe não for daltónica nem portadora (DD) e o pai possuir visão normal (D), nenhum dos descendentes será daltónico nem portador.
Se a mãe possuir visão normal (DD) e o pai for daltónico (d), nenhum dos descendentes será daltónico, porém as filhas serão portadoras do gene (Dd).
Se a mãe for portadora do gene (Dd) e o pai possuir visão normal (D), há a probabilidade de 50% dos filhos serem daltónicos e 50% das filhas serem portadoras do gene.
Se a mãe for portadora do gene (Dd) e o pai for daltónico (d), 50% dos filhos e das filhas serão daltónicos.
Se a mãe for daltónica (dd) e o pai possuir visão normal (D), todos os filhos serão daltónicos (d) e todas as filhas serão portadoras (Dd).
Se a mãe for daltónica (dd) e o pai também (d) 100% dos filhos e filhas também serão daltónicos.
O pai só transmite o daltonismo às filhas.
Hemofilia
É uma doença genética em que a capacidade de coagulação do sangue é muito reduzida por falta de um fator de coagulação presente no plasma.
Há vários tipos de hemofilia (A,B e C), a mais frequente é a A e é causada pela falta de um fator (VIII) de coagulação que é controlado por um gene existente no cromossoma X. A hemofilia B é também causada por falta de um fator (IX), também sintetizado pelo cromossoma X, e a C está ausente o fator XI mas sintetizado por um gene autossómico.
A doença pode ser controlada administrando o fator que está ausente no sangue.
A herança da hemofilia A segue o mesmo padrão do daltonismo.
Se um homem com hemofilia A (XhY) se casar com uma mulher não hemofílica (XHXH) nenhum de seus filhos e filhas terá a doença, mas as filhas serão portadoras e poderão ter filhos hemofílicos, mesmo que se casem com homens normais.
Nas árvores genealógicas utiliza-se o h para designar o gene responsável pela hemofilia e o H para designar o gene responsável pela normalidade do indivíduo.

http://biologia-ap.no.comunidades.net/index.php?pagina=1196451636
Portanto, a hemofilia (e outras ligadas ao sexo) transfere-se de um homem para o seu neto por meio de sua filha.
As mulheres hemofílicas são muito raras (XhXh), pois é necessário que um homem hemofílico (XhY) se case com uma mulher portadora (XHXh).
1.2. Organização e regulação do material genético
![]()
Organização do material genético
O material genético encontra-se nos seres eucariontes no núcleo, e está organizado em cromossomas/cromatina. Ao conjunto dos cromossomas da célula dá-se o nome de cariótipo. Mas há também material genérico extranuclear, nomeadamente nas mitocôndrias e cloroplastos que possuem as suas próprias moléculas de DNA. Estas moléculas, DNA mitocondrial e plastidial, não se encontra associadas às proteínas (histonas) como o DNA nuclear. Estas moléculas apresentam os seus próprios genes que estão diretamente relacionado com as funções dos respetivos organitos, o que lhes confere alguma autonomia, no entanto, o DNA nuclear também contém os genes para o metabolismos destes organitos, por isso a autonomia não é total.
Os cromossomas destes organitos são circulares.


Como já sabes o DNA nuclear constitui uma dupla hélice e codifica cerca de 100.000 genes, o DNA mitocondrial representa apenas 1 a 2% do DNA celular, em duplo filamento circular, e codifica cerca de 37 genes. Estes genes estão associados à produção de energia. Se existirem mutações nestes genes pode haver uma baixa produção de ATP.
Como são transmitidos aos descendentes?
Já vimos na reprodução que na fecundação, o espermatozoide tem pouco citoplasma, pelo que na formação do zigoto, o espermatozoide não contribui com o citoplasma. Por esta razão, as mitocôndrias do descendente, vêm todas da mãe.
Influência de agentes endógenos e exógenos na expressão génica
Na maioria dos casos existe correspondência entre o genótipo e o fenótipo expresso, quer dizer que os genes relativos ao fenótipo se expressaram a 100%, diz-se nestes casos que estes genes têm uma penetrância. Mas é na maioria dos casos, numa minoria não existe esta correspondência total de expressividade de um gene. Esta expressividade pode ser influenciada por diversos fatores, exógenos, por exemplo o ambiente influencia a cor da pele, e endógenos, por exemplo as hormonas, ou a existência de genes letais ou supressores.
Como também já vimos, e um exemplo é a cor de olhos, determinadas características são determinadas não apenas por um gene mas por vários genes influenciados pelo ambiente. Chamam-se a estas caraterísticas, características quantitativas (determinadas por vários genes e influenciadas pelo ambiente).
![]() Regulação da expressão génica
Regulação da expressão génica
Todas as células de um organismo contêm o mesmo conteúdo genético. O que muda entre células, são os genes que são expressos, ou seja o complemento de proteínas que cada célula produz que está diretamente relacionado com a especificidade da célula que constitui um tecido. As células não expressam todos os seus genes ao mesmo tempo, a forma como elas induzem ou reprimem os seus genes é que as faz diferenciadas.
Regulação da expressão génica dos procariontes
Os seres procariontes expressam os seus genes de forma diferente dos eucariontes, nestes é mais complexo e porque têm uma maior quantidade de informação genética, a células eucariótica é compartimentada podendo a regulação dos genes se dar em diferentes locais e também porque os eucariontes são seres multicelulares.
A regulação génica está relacionada com a eficiência energética e os consumos de recursos disponíveis. Os organismos ajustam o seu metabolismo às modificações que ocorrem no meio.
Nos seres procariontes, as bactérias, a quantidade de proteínas/enzimas é muito variável, está relacionada com as suas necessidades. A regulação, nestes seres é feita através: de mecanismos que impedem a transcrição de genes que codificam as proteínas que a célula não está utilizar e outros que induzem a transcrição das proteínas que estão a ser necessárias ao metabolismo da célula; da inativação do mRNA, antes de ser traduzido; do impedimento da tradução do mRNA nos ribossomas e da inativação da proteína que foi produzida e não é necessária impedindo-a de funcionar.
Nos seres procariontes os genes que codificam proteínas envolvidas no mesmo processo metabólico localizam-se no DNA de uma forma contígua e são controlados por uma unidade estrutural, o operão. Todos os genes estruturais (porções de DNA que codificam proteínas de uma mesma via metabólica), são transcritos por um único mRNA.

Estrutura do operão (opéron)
É constituído por um grupo de genes ordenados lado a lado que codificam enzimas que trabalham numa mesma via metabólica, controladas por um único promotor.
- região do promotor, porção do DNA à qual se liga a RNA polimerase, dando inicio à transcrição;
-região do operador- local entre o promotor e os genes estruturais e onde por vezes se liga o repressor (proteína especial que quando ligada ao operador impede que a RNA polimerase não chegue aos genes estruturais impedindo a transcrição);
-dois ou mais genes estruturais;
- e no início do operador existe um gene regulador ou repressor que é responsável pela produção do repressor que se ligará, como já foi referido, ao operador. Embora o gene regulador afete o operão não é considerado parte do operão.
A proteína reguladora pode ser sintetizada em duas formas: ativa ou inativa. Dependendo do estado no qual ela é sintetizada teremos os dois tipos básicos de funcionamento do operões: os operões de indução e os operões de repressão. Os de indução estão, normalmente associados a processos catabólicos (ex: metabolismo do glúcidos) e os de repressão a processos de anabolismos (síntese de aminoácidos).
http://aprendendogenetica.blogspot.pt/2011/05/aulas-6-e-7-biologia-regulacao-da.html
https://www.youtube.com/watch?v=lK4LadkOll0
![]() Em
1965, dois investigadores franceses, François Jacob e Jacques Monod, trabalharam
com a bactéria Escherichio coli e os resultados do estudo valeram-lhe o
prémio Nobel pelo contributo que deram à ciência sobre a expressividade dos
genes. O estudo visou sobre o metabolismo da lactose (operão Lac) e a síntese do
triptofano (operão trp).
Em
1965, dois investigadores franceses, François Jacob e Jacques Monod, trabalharam
com a bactéria Escherichio coli e os resultados do estudo valeram-lhe o
prémio Nobel pelo contributo que deram à ciência sobre a expressividade dos
genes. O estudo visou sobre o metabolismo da lactose (operão Lac) e a síntese do
triptofano (operão trp).

http://www.theguardian.com/science/2013/apr/25/francois-jacob
O Operão da lactose é comandada por 3 genes estruturais, a LacZ, LacY e LacA, que codificam as enzimas necessárias ao metabolismo da lactose, tem ainda o gene operador, segmentos do DNA onde se pode ligar o repressor); um gene promotor, segmento do DNA onde se liga a RNA polimerase desde que o promotor esteja livre do repressor; e ainda o gene regulador, gene responsável pela produção do repressor e que se localiza fora do operão.
Na ausência de lactose
O gene regulador determina a síntese do repressor que está ativo. O repressor liga-se ao operador, a enzima RNA-polimerase não se liga ao promotor, e bloqueia a transcrição dos genes estruturais (Z,Y,A) que codificam as 3 enzimas, portanto não se formam, porque não há necessidade.
Na presença de lactose
É um mecanismo indutivo pois a presença da lactose promove o funcionamento dos genes estruturais (X,Y,A). O gene regulador determina a síntese do repressor e a lactose liga-se a este, inativando-o. O gene operador fica desbloqueado e a enzima RNA-polimerase liga-se ao promotor, os genes estruturais são transcritos e ocorre a síntese das enzimas.
O operão lactose é um mecanismo do tipo indutivo dado que é a presença da lactose que induz o funcionamento dos genes estruturais.


O operão triptofano é um mecanismo do tipo correpressor
Nos operões de repressão, a proteína reguladora é produzida na forma inativa, ou seja, incapaz de reconhecer e de se ligar ao operador. Neste caso, a proteína reguladora somente ganha a capacidade de se ligar ao operador em presença de uma molécula coadjuvante, denominada correpressor.
Desta forma temos duas situações:
-sem a presença do correpressor, a proteína reguladora não se liga ao operador e a RNA polimerase realiza a transcrição
-ou na presença do correpressor, a proteína reguladora ganha afinidade pelo operador, impedindo que a RNA polimerase realize a transcrição.
O triptofano é um aminoácido que pode ser produzido pela E.coli ou vir do meio externo e se isto acontecer a bactéria não necessita de sintetiza-lo, logo também não é necessário produzir enzimas que são necessárias à sua síntese. Este mecanismo, é como foi referido, repressivo. O triptofano vindo do exterior liga-se ao operador e impede a transcrição dos genes estruturais, responsáveis pela produção de enzimas envolvidas na síntese do triptofano. Chama-se ao triptofano de correpressor.
Na ausência de triptofano, o gene regulador produz um repressor que é inativo, o gene operador fica livre, a RNA-polimerase liga-se ao gene promotor, dá-se então a transcrição e a consequente síntese de enzimas necessárias à síntese do triptofano.


Um regulador pode controlar um grupo de operões e constitui-se um regulão.
Por exemplo: operões com intervenção no catabolismo de glícidos são controlados em simultâneo pelo mesmo gene regulador, tornando mais eficaz e rápida a conversão de glícidos em glicose.
Nas células procarióticas (como é o caso da bactéria Escherichia coli), a ativação ou repressão dos operões resulta muitas vezes de alterações ambientais, tais como variações bruscas de temperatura, alterações na quantidade de água, de oxigénio e de outras substâncias presentes no meio como por exemplo a lactose ou o triptofano.
2. Alterações do material genético
![]()
2.1. Mutações
O genoma dos indivíduos experimenta, em várias circunstâncias, alterações, alterações estas que podem ser diárias, no entanto a capacidade para reparar alterações por parte das células é extraordinária e a grande maioria desaparece, restando apenas algumas anomalias. As alterações permanentes no DNA da células designam-se por mutações e os indivíduos que as possuem, por mutantes. Estas mutações podem afetar um único gene ou afetar a globalidade dos cromossomas do individuo.
Os efeitos de uma mutação numa célula são imprevisíveis, por vezes podem ser benéficas, outras vezes prejudiciais, podendo inclusive levar à morte e outras vezes o efeito pode ser neutro.
As mutações podem ocorrer nas células somáticas ou nas linhas germinativas e têm consequências diferentes. Uma célula somática que sofreu uma mutação, por meiose, cria clones que poderá afetar a vida do indivíduo mas não a dos seus descentes e as mutações na células das linhas germinativas pode ser transmitida aos seus descendentes.
Podemos ter diferentes tipos de mutação dependentes do local e da fase onde ocorrem:
Mutações génicas, quando são mutações ao nível dos nucleótidos de um gene;
Exemplo a anemia falciforme (substituição de um nucleótido por outro).
Mutações cromossómicas, quando as mutações afetam grandes porções do genoma (exemplo na meiose e mitose), partes de cromossomas ou cromossomas inteiros;
Mutações cromossómicas estruturais, em que há quebra de cromossomas e a união e reorganização, alterando o DNA;
Tipos de alteração cromossómica estruturais:
Deleção
Uma parte do cromossoma eliminado, o que implica a perda de muitos genes. Ocorrem durante o emparelhamento dos cromossomas na meiose.
Um exemplo humano é o síndrome do miado do gato, em que falta um fragmento do braço curto do cromossomo 5. Este síndrome carateriza-se por atraso mental, microcefalia, aspeto arredondado da face e de choro semelhante a um miado de gato.

Inversão
Uma parte do cromossoma quebra-se e sofre rotação de 180º e solda-se novamente em posição invertida sem alterar a sua posição no cromossoma.
Translocação
Trata-se da troca de partes de cromossomos não homólogos, é diferente do que ocorre no crossing-over.
É possível que a translocação tenha sido um mecanismo de formação de novas espécies. Há quem coloque a hipótese sobre algumas espécies de drosófilas, em que o número de cromossomas é diferente, podendo ter sido originado por ancestral comum que tenha sofrido translocações de diversos tipos.
Duplicação
Na duplicação, há a formação de um segmento adicional num dos cromossomas, ou seja, há duplicação. De modo geral, as consequências de uma duplicação são bem toleradas pois não há falta de material genético.

Mutações cromossómicas numéricas, em que há uma não disjunção de cromossomas homólogos na meiose I ou disjunção de cromatídeos na meiose II e o resultado final é um número diferente de cromossomas. A maior parte dos embriões abortam espontaneamente.
Diz-se euploidia quando o cariótipo apresente o número normal de cromossomas (2n=46) e Aneuplodia quando apresenta ou excesso ou falta de cromossomas do cariótipo da espécie.
Diz-se Trissomia – quando apresenta três cromossomas em vez de um único par de cromossomas homólogos (2n+1); Monossomia - só apresenta um cromossoma em vez de um par de homólogos (2n-1); Nulissomia – caso mais raro, em que pode não existir nenhum cromossoma de um determinado par (2n-2).

A trissomia 21 ou síndrome de Down é um exemplo destas mutações. Há uma cópia a mais do cromossoma 21.

Síndromes resultantes dos cromossomas sexuais
Síndrome de Turner - deficiência de um cromossoma X nas mulheres (45,X). É a única monossomia viável na nossa espécie: Perda de um cromossoma (2n -1). Esta anomalia é letal para o sexo masculino (aborto espontâneo). Mulheres pequenas sempre com ar infantil, não têm carateres sexuais secundários e os ovários não funcionam.

Síndrome de Klinefelter - É também uma trissomia mas esta ligada ao sexo masculino: Acréscimo de um cromossoma (2n+1)- acréscimo de um cromossoma X nos homens ( 47,XXY) e nas mulheres (47, XXX). Nas mulheres este tipo de trissomia não origina nada de especial, para além de serem mulheres altas, no homem, são altos, normalmente estéreis.

Poliploidia
A Poliploidia é a ocorrência de dois ou mais pares de cromossomos homólogos numa célula. O número de cromossomas destes seres é um múltiplo do número de cromossomas do gameta: 3n, 4n…
Pode resultar da não disjunção dos cromossomas homólogos na mitose ou na meiose, pode resultar de não haver citocinese, ficando a célula com 4n.
Estes seres não se podem cruzar com os da sua espécie mas podem reproduzir-se assexuadamente. É muito frequente nos vegetais e pode trazer vantagens para o homem, que provoca a polipoidia das plantas em laboratório para seu belo proveito.
Um exemplo de polipoidia é o facto de muitas plantas que apresentam esta mutação, poderem apresentar frutos muito grandes para lhes permitir a colonização rápida noutras áreas.


http://www.netxplica.com/diapositivos/biologia12/poliploidia/19.html
![]() Mutações e o desenvolvimento biotecnológico
Mutações e o desenvolvimento biotecnológico
Qualquer agente que seja responsável por uma mutação é um agente mutagénico e o processo que leva ao seu aparecimento é a mutagénese. Embora as nossas células tenham grande capacidade de reparação das alterações, muitas vezes não conseguem.
As mutações podem ser espontâneas ou induzidas, ambas são alterações definitivas no DNA da célula, mas a espontânea está relacionada com o mau funcionamento da células e as induzidas têm causas externas, os agentes mutagénicos.
As espontâneas podem ser causadas pela forma incorreta que se ligam as bases, por exemplo em vez da citosina emparelhar com a guanina, emparelha com a adenina; por vezes reações químicas que ocorrem na célula podem gerar alteações nas bases e até a DNA polimerase pode gerar erros na duplicação do DNA.
Nas induzidas que se devem a causas externas os agentes mutagénicos podem ser físicos, como as radiações ionizantes (raio X, radiações alfa, beta e gama) e radiação ultravioleta; e podem ser químicos como alguns corantes e conservantes, benzeno, benzopireno, alcatrão do fumo do cigarro, etc.
As mutações genéticas e o Cancro, tumor maligno ou neoplasia maligna
O cancro é uma doença que apresenta o crescimento de um tecido neoformado. Estima-se que 25% dos europeus, numa fase da vida, apresente esta doença. Têm origem genética e apenas uma minoria é que são hereditários, estes manifestam-se, normalmente em forma de multitumores, a grande maioria são esporádicos (95%) e nestes se incluem a ação de determinados vírus e de substâncias tóxicas.
As células cancerígenas (massa de células neoformadas) distinguem-se das que lhe deram origem por:
-Possuírem núcleos de forma e tamanho diferentes;
-Apresentarem alterações dos mecanismos que regulam a divisão celular (alterações devido a um aumento da estimulação da divisão celular ou devido a deficiências nos mecanismos que impedem a divisão celular);
-Apresentarem alterações dos mecanismos que regulam a apoptose;
- Terem a capacidade de invadir outros tecidos através das metástases, aproveitando a corrente sanguínea e linfática.
Etapas de formação de metástases: estas células têm a capacidade de produzir substâncias químicas desenvolver vasos sanguíneos que fornecem oxigénio ao tecido neoformado (angiogénse) e produzem enzimas digestivas que destroem as células vizinhas e introduzem-se no sangue e na linfa, indo instalar-se noutros tecidos.
Um ser multicelular é o resultado do equilíbrio entre as numerosas células que num determinado momento se dividem (proliferação celular) e noutro se autodestroem numa morte programada (apoptose).
A apoptose, é então, o conjunto de processos genéticos programados que levam à morte da célula, desejável e necessária que participa na formação dos órgãos e que persiste em alguns sistemas adultos como a pele e o sistema imunológico.
Existe ainda um outro processo que leva à morte da célula, a necrose, mas este deve-se à falta de nutrientes essenciais ou à ação de agentes tóxicos na célula.
Diferenças entre a necrose e a apoptose
Na apoptose, microscopicamente ocorre fragmentação nuclear e celular em vesículas apoptóticas.
A célula condensa-se, encolhe, e começam a formar-se bolhas, a cromatina é compactada, formando massas concentradas na parte interna do núcleo, que se parte, levando à formação das vesículas apoptóticas.
A apoptose é diferente da necrose, não existe libertação do conteúdo celular para o interstício e portanto não se observa inflamação ao redor da célula morta.
A necrose difere da apoptose por representar um fenómeno degenerativo irreversível, causado por uma agressão intensa. Trata-se pois da degradação progressiva das estruturas celulares sempre que existam agressões ambientais severas.

http://www.virtual.unifesp.br/unifesp/bio40/apoptose/
http://www.noas.com.br/ensino-medio/biologia/citologia/apoptose-e-necrose/ animação
Diariamente o nosso organismo debate-se com células neoplásicas que são eliminadas por apoptose. Se estas células não forem destruídas, porque foram geneticamente alteradas, surge o cancro.
Os cientistas identificaram dois tipos de genes, quando mutados, podem causar cancro:
- os proto-oncogenes ou genes promotores de crescimento, com a capacidade de estimular a divisão celular encontrando-se no estado inativo em células que não estão no estado de proliferação. A ativação deste gene por mutação transforma-se num oncogene e este pode provocar um crescimento celular descontrolado, causando a formação de um cancro. A existência de um gene mutado num alelo é suficiente para desenvolver-se o cancro.
- os genes supressores de tumor (tumorais), regulam o ciclo celular, produzindo proteínas que bloqueiam determinadas fases do ciclo. Estes genes contrabalançam o estímulo proliferativo dos proto oncogenes. Quando ocorre uma mutação neste gene, deixam de prevenir a multiplicação descontrolada das células. Ao contrário dos oncogenes, para que se desenvolva o cancro, é necessário a existência da mutação dos dois alelos.
As células do nosso corpo são, assim, reguladas de forma a que haja um balanço entre os genes que induzem o crescimento celular (proto-oncogenes) e os genes que bloqueiam o crescimento ( supressores de tumores).
As mutações têm aspetos positivos e negativos, como já vimos, se por um lado podem originar doenças letais, ou acarretar anomalias graves, por outro, são responsáveis pelo aumento da variabilidade genética, promovendo a evolução das espécies e ao nível do homem, o conhecimento de genética tem permitido à nossa sociedade utilizá-las, como por exemplo, na criação de determinadas mutações em seres vivos que consumimos. É sabido que ingerimos muitos alimentos cujo o cariotipo foi alterado com vista a uma maior rentabilidade, ou na introdução de determinadas características mais apreciadas pelo homem.
Os prós da biotecnologia… e os contra? Pesquisar.
![]() 2.2. Fundamentos da engenharia genética
2.2. Fundamentos da engenharia genética
A engenharia genética, é uma ciência recente, possibilita a manipulação de genes de um determinado organismo, geralmente de forma artificial, através de técnicas.
Com o objetivo de produzir organismos geneticamente melhorados para desempenharem melhor suas funções e produzir substâncias úteis, através de técnicas de duplicação, transferência e isolamento de genes.
As técnicas são os processos que a própria célula utiliza, mas recriadas em laboratório.
Enzimas de restrição
A grande descoberta das enzimas de restrição originou uma revolução ao nível das ferramentas da engenharia genética.
As enzimas de restrição cortam o segmento em dupla hélice de DNA em locais específicos. Mas a enzima só cortará a molécula se encontrar uma determinada sequência de bases nas duas hélices lidas sempre de 5´para 3´. Sempre que encontrar essa sequência a enzima corta o DNA, originando pequenos fragmentos de DNA enrolados em dupla hélice e que terminam em pequenas porções de DNA em cadeia simples (extremidades coesivas). Estas extremidades, podem ligar-se por complementaridade a outras moléculas de DNA , e esta ligação tem a intervenção de outras enzimas, as ligases do DNA. Foi esta descoberta que permitiu aos cientistas transferirem o DNA de um ser para outro, a que se dá o nome de vetor (o ser que recebe o DNA). Os vetores que são utilizado na engenharia genética, são normalmente, as bactérias, mas também se utilizam vírus, leveduras e outras células eucarióticas. As bactérias contêm pequenas moléculas circulares de DNA, designadas por plasmídeos.
“O plasmídio é o material genético circular não ligado ao cromossoma que fica espalhado pelo hialoplasma das bactérias. Ele sofre o mesmo processo do DNA cromossomal de transcrição e tradução, além de, se multiplicar a cada divisão celular, passando uma cópia para cada célula “filha”.”

http://www.sobiologia.com.br/conteudos/Biotecnologia/recombinante.php
O plasmídeo é retirado da bactéria para que se possa introduzir o DNA do outro organismo.

Para entendermos melhor o processo vamos ver passo a passo:
|
http://www.sobiologia.com.br/conteudos/Biotecnologia/recombinante.php |
Quer-se estudar um gene humano que produz uma proteína
que não se sabe a função.
|

DNA recombinante
A engenharia genética recorre a técnicas de biologia molecular para a manipulação de genes, alterando a sua estrutura de DNA as suas características. Esta técnica é conhecida por DNA recombinante e permite recombinar o material genético de um gene de um determinado individuo, com material genético de uma outra espécie. Esta técnica é utilizada para: aumento de produtividade de determinadas plantas; aumento nutritivo de determinada espécie; aperfeiçoamento e rentabilização dos processos de produção de vinho, iogurtes, queijos; em testes de paternidade e diagnósticos de doenças infeciosas e genéticas; desenvolvimento de organismos geneticamente modificados (OGM); estudos de mecanismos de replicação e expressão génica; determinação da sequência de um gene e proteína codificada; produção de medicamentos, entre outras aplicações.
Um dos primeiros medicamentos feito através da DNA recombinante foi a insulina humana.

http://www.medicinageriatrica.com.br/2013/01/16/terapia-genica-producao-de-insulina/
Produção de insulina humana
http://pt.scribd.com/doc/46819023/APLICACOES-DA-ENGENHARIA-GENETICA-Resumo
http://www.medicinageriatrica.com.br/2013/01/16/terapia-genica-producao-de-insulina/
DNA complementar (cDNA)
O DNA complementar (cDNA) é um outro processo utilizado, na maior parte das vezes, como um auxiliar da técnica do DNA recombinante.
O cDNA é obtido a partir do RNA mensageiro (rRNA) maturado (já não possui intrões) por complementaridade de bases. Esta transcrição em sentido inverso é conseguida através da ação da enzima viral denominada transcriptase reversa. Assim, o mRNA funciona como molde para a síntese de uma cadeia de DNA. Após a formação da cadeia de DNA, a enzima DNA polimerase atua, formando, por complementaridade, a outra cadeia de DNA a partir dos nucleótidos presentes no meio, constituindo-se uma molécula estável.

Porto editora
Reação de Polimerização em Cadeia (Polymerase Chain Reaction - PCR)
Este processo baseia-se no processo de replicação de DNA que ocorre in vivo e é uma das técnicas para clonar DNA de modo a obter grandes quantidades de uma determinada porção de DNA, a partir de uma pequena amostra.
Em primeiro lugar, deve-se extrair o material genético da célula com interesse sem o danificar. Normalmente, o material extraído é o DNA mas pode-se trabalhar também com o RNA num método designado RT-PCR (reverse transcription polymerase chain reaction) que é uma variante do PCR e possui outras aplicações.
Depois de extraído o DNA, é aquecido o DNA para a separação das duas cadeias, é-lhe adicionada uma mistura que contém nucleótidos (adenina, timina, guanina e citosina), nucleótidos iniciadores (primers) e a enzima DNA polimerase. Toda esta mistura é colocada na máquina de PCR, o termociclador, que efetua ciclos de temperatura preestabelecidos com tempos exatos. Repete-se o procedimento até se obter as cópias pretendidas.
A polimerização em cadeia faz rápidas replicações de determinadas porções de DNA.
 Termociclador
Termociclador